Mobility and orientation mechanism of the Cucurbita maxima

In this delivery, I will share with the whole community of the HIVE platform, especially with the members of the academic community #STEMsocial, content about the PHYTOMECHANICAL ABILITY that plants exhibit, specifically aspects related to the promotion mechanisms used by plant species. In this opportunity we will outline the phytomechanical model of mobility and orientation that the Cucurbita maxima has, associating eco-physiological responses by phototropism and tigmotropism.
Introduction
Unlike animals, plant species lack a system of movement by muscular action, whose orientation, length and speed are controlled by neurons that intervene physiologically in the execution of each movement. On the contrary, plant species have developed a series of morphological adaptations as a result of evolution, in order to generate biomechanical responses to external stimuli [1].
With regard to mobility and orientation, plant species have adapted morphological structures such as trichomes, a vegetative organ capable of perceiving environmental stimuli and inducing phytomechanical responses that guarantee growth, development and production in plants [2].
Therefore, the purpose of this post is to describe the phytomechanical model that Cucurbita maxima has, in order to guarantee the mobility and orientation of its vegetative structures.
Phyto-mechanical ability
As it has been indicated in the previous posts, the plant species possess phytomechanical ability to carry out certain movements, that according to the executed speed can be; fast, slow, prolonged or short, these movements have been categorized as tropisms and nasties, both biological responses are related to the asymmetric growth of certain organographic structures in the plants, that when perceiving external stimuli of variable intensity, as for example photons of light, they reorient their topographic location in the plant [3].
Physiologically, this capacity that organographic structures have, to reorient their topographical location in the plant, is attributed to certain vegetative elements such as trichomes, which, depending on the wavelength and light intensity captured, generate a strong histological tension, mainly colenquimatic, at the base of petioles and vegetative tendrils in order to control the orientation of the leaf blades (leaves) and thus have a greater photoreceptor area.
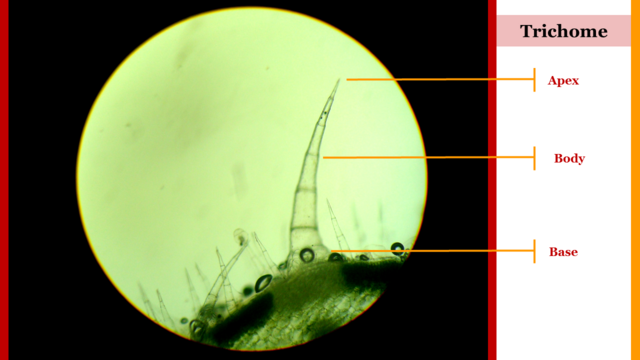
Fig. 2. Cucurbita maxima pluricellular trichome. Author: @lupafilotaxia.
Phototropism and tigmotropism are movements exercised by plant organographic structures, the first as a mechanical response sensitive to sunlight, and the second as a biomechanical action mediated by stimuli from external contacts, it is normal to detect these movements in specific plant groups, however, it is not common to observe both responses associated with the same plant structure [4].
Associated biomechanical structures
The tendrils and trichomes, can act as associated biomechanical devices, generating phytomechanical responses in certain plant species, whose biological mechanism works as follows; the trichomes perceive the wavelengths and orient the tendrils towards external supports, which are in the same direction of light interception [2].

Fig. 3 Circumnutated tendril with trichomes, seen in apical stem segment in Cucurbita maxima. Author: @lupafilotaxia.
Morphology of the phytomechanical organ
The manuscript deals with two biomechanical structures. The main phytomechanical organ considered in the post is the trichome, a botanical element of variable morphology that protrudes from the epidermal tissue, of unicellular or pluricellular constitution and of multiple functions, among which the most important are; adsorption, perception, protection and secretion.
The second phytomechanical organ considered is the tendril, a botanical structure with a filamentous morphology, with touch-sensitive areas in the apex region, it has epidermal cells with cell walls with a high content of cellulose and hemicellulose that give it rigidity, it also has abundant cholenchymatic cells (mechanical tissue) in the whole periphery with the capacity to roll up.

Fig. 4 Biomechanical action of tendril in circumnutation on leaf blade in species of taxon poaceae, response mediated by light perception captured by trichomes in Cucurbita maxima. Author: @lupafilotaxia.
Both organographic structures show biomechanical behavior, mediated by genetic information at the cellular level, which gives them the ability to perceive and transmit movements when receiving stimuli.
Climbing mechanism
Field phytomechanical study
In order to elucidate the phytomechanical model of mobility and orientation that the Cucurbita maxima has, associating eco-physiological responses by phototropism and tigmotropism, a field trip was carried out in the cultivation area of the UNESUR Botanical Garden located in the biogeographical zone of the South of Lake Maracaibo, Zulia State - Venezuela, with the aim of describing the biomechanical strategies of ascent physiologically executed by the species.
Mechanical climbing strategies
Phototropism and tigmotropism are the systems of mobility and orientation exhibited by Cucurbita maxima (Auyama), as mechanical strategies to ascend and capture sunlight. These biological abilities are executed through the histological tension triggered in the cholenchymatic cells found at the base, body and apex of the tendrils, generating movements by circumnutation on external structures, which are in the same direction of the light interception perceived by the trichomes.
Strategy I: Trichomes as light sensors
In order to take advantage of the photons light, the trichomes of Cucurbita maxima, when perceiving the wavelengths send traction signals to the colenquimatic tissue (mechanical cells) located in the base, body and apex of the tendrils as a biomechanical action for the reorientation of the foliar structures towards the captured sunlight [2].
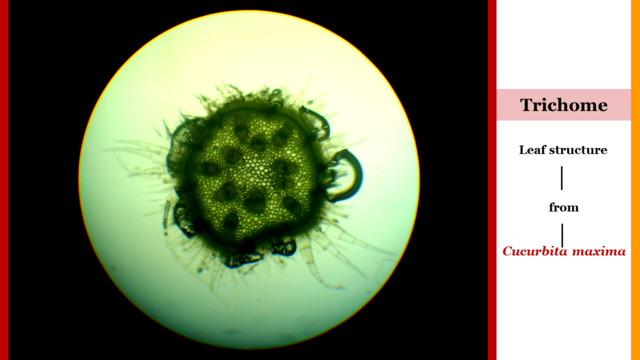
Fig. 5 Transversal view of multicellular trichomes located in the organographic structure of a Cucurbita maxima. Author: @lupafilotaxia.
Strategy II: Earring with an upright apex
The Cucurbita maxima, has a similar mechanism of mobility and circumnutation as the one observed in Passiflora edulis (Parchita), therefore in a similar way the meristematic tissues multiply and generate a foliar aggregate called tendril, which tends to develop longitudinally and in an erect way in search of external contacts.

Fig. 6 Vegetative tendrils of Cucurbita maxima, a tendril with an upright and curved apex can be seen. Author: @lupafilotaxia.
Strategy III: Curved Apex Earring
The contact of the apex of the earrings with external elements triggers the curvature of the earring, a biomechanical strategy used to create a kind of hook to hold on to.
Strategy IV: Earring with circumnutation
After the contact of the Cucurbita maxima tendrils with some external element, a coil is initiated by circumnutation in a helical way, with the lower, middle and upper segments of the external support, until guaranteeing an area rich in light photons that favours the photosynthetic activity.

Fig. 7 Cucurbita maxima earrings, with lower segment circumnutation. Author: @lupafilotaxia.

Fig. 8 Cucurbita maxima earrings, with medium segment circumnutation. Author: @lupafilotaxia.

Fig. 9 Cucurbita maxima earrings, with circumnutation in upper segment. Author: @lupafilotaxia.
Indicators of phytomechanical tropism
To describe the indicators of mobility and orientation, by means of phototropism and positive tigmotropism in Cucurbita maxima, the distance was measured in the different biomechanical events observed in situ.
Register 1
The tendrils were observed in circumnutation condition, under the leaf blades of Cucurbita maxima and in the lower zone of the intercepted external element at a distance of approximately 5 cm from the soil.
Register 2
They were observed under circumnutation conditions, on dry intercepted vegetative structures of species of the taxon poaceae, at a distance of approximately 10 cm from the ground.

Fig. 10 On the left, we can see Cucurbita maxima tendrils, in circumnutation on dry poaceae leaf tissue located 5 cm aprox. from the soil, on the right, circumnutation on dry poaceae leaf tissue located 10 cm aprox. from the soil. Author: @lupafilotaxia.
Register 3
The last record made in situ, was taken at 15 cm height, over fresh vegetative structures of intercepted species of the taxon poaceae, in an area totally cleared for the capture of sunlight.

Fig. 11 Cucurbita maxima tendrils are observed, in circumnutation on fresh poaceae leaf tissue located 15 cm aprox. from the soil. Author: @lupafilotaxia.
SCIENTIFIC CONTRIBUTIONS OF THIS PUBLICATION
- The content and considerations generated in the post, expand the theoretical-scientific vision on the biomechanical strategies of mobility and orientation, through responses by phototropism and tigmotropism observed in the plant species Cucurbita maxima, by responding effectively to the capture of light wavelengths available in their ecophysiological environment, through the use of morphological structures as phytomechanical resources, which constitute an important factor and guarantee optimal levels of photosynthetic activity. Within these biomechanical structures, trichomes and tendrils are identified, which are vegetative elements that, as a result of evolution and genetic adaptation, allow C. maxima, intercept the different intensities of light, and generate biological responses in the cholenchymatic cells, thus orienting the growth of the organs in the plant.
BIBLIOGRAPHICAL REFERENCES CONSULTED AND CITED:
[1] Lambers H., Chapin F., and Pons T. Plant Physiological Ecology. Springer, Nueva York, EUA. 1998;540. Article: Online access
[2] Zaja U., Kucharski S., and Guzek D. Are trichomes involved in the biomechanical systems of Cucurbita leaf petioles?. Planta. This article is published with open access at Springerlink.com. 2015;13. Article: Online access
[3] Hohm T., Preuten T. and Fankhauser C. Phototropism: Translating light into directional growth. American Journal of Botany. 2013;100;1:47-59. Article: Online access
[4] Domínguez E., Cuartero J., and Heredia A. An overview on plant cuticle biomechanics. Plant Sci. 2011;181:77-84. Article: Online access
[5] Briggs W., and Olney M. Photoreceptorsin plant photomorphogenesis to date: Five phytochromes, two cryptochromes, one phototropin, and one superchrome. Plant Physiology. 2001;125:85-88. Article: Online access
[6] Leroux O. Collenchyma: a versatile mechanical tissue with dynamic cell walls. Ann Bot. 2012;110:1083-1098. Article: Online access
[7] Meisel L., Urbina D., y Pinto M. Fotorreceptores y respuestas de plantas a señales lumínicas. Fisiología Vegetal. Capitulo XVIII. 2011;18. Article: Online access
ATTENTION
Readers and followers
If you wish to read more scientific articles in English or Spanish, of excellent academic quality, do not hesitate to visit #STEMSocial and #STEM-espanol, communities that promote scientific content mainly in the areas of Science, Technology, Engineering and Mathematics.

0
0
0.000
#Posh Twitter:
https://twitter.com/lupafilotaxia/status/1260030961908985860
Thanks for your contribution to the STEMsocial community. Feel free to join us on discord to get to know the rest of us!
Please consider supporting our funding proposal, approving our witness (@stem.witness) or delegating to the @steemstem account (for some ROI).
Thanks for using the STEMsocial app and including @stemsocial as a beneficiary, which give you stronger support.
Fantastic photographs and excellent schematic explanation of the Cucurbita maxima, a material worthy of stemsocial. Thank you for this!!! professor @lupafilotaxia
Greetings @carloserp-2000, thanks for the positive comments, socializing the phytomechanical responses of the plants, is a very exciting point for any botanist.
I agree with @carloserp-2000! The pictures are amazing and illustrate nicely the text. On my side, I will never look at the squash growing at home in the same way (they features things similar to what is exposed here, if I am not mistaking myself ;)
Greetings @lemouth, thank you for visiting the blog. In fact, our way of observing plants changes when we manage to interpret the physiological responses involved in the movements and survival strategies employed by plants.
The plants you have in your house probably belong to the same biological taxon. However, Cucurbita maxima receives multiple common names, in the case of South America the most common are Auyama, Calabacin, or Zapallo, while in North America and Europe, it is identified by Calabaza, however, although in most cases it is of the same genus (Cucurbita), this does not mean that it is the same species, since Cucurbita maxima is often confused with Cucurbita pepo, and Cucurbita moschata.
Thanks for the clarifications. So probably the same class of species, but not more. Is that correct?
Right, to the same taxon at the gender level, i.e., the Cucurbita group.
If the plants in your house have similar characteristics to those described in this post, their taxonomic location is something like this:
Kingdom: Plant
Division: Magnoliophyta
Class: Magnoliopside
The family: Cucurbitaceae
Gender: Cucurbita
Species: ?
To identify the species, you can send photos of the leaves and fruits.
I will do that tomorrow. Unfortunately, they are not in a great shape... They got some rain and seem to struggle since then... :/